The Systemic Consequences of STAT3 Dysregulation in Long COVID:
- Graham Exelby
- Jun 16, 2025
- 6 min read

Updated: Sep 28, 2025
A Unifying Molecular Pathway Across Malignancy, Autoimmunity, and Immune Dysregulation
Dr Graham Exelby June 2025- revised September 2025
Abstract
Long COVID is increasingly recognized as a condition marked by chronic immune–metabolic dysregulation, with Signal Transducer and Activator of Transcription 3 (STAT3) emerging as a pivotal amplifier within interconnected inflammatory circuits. STAT3 activation is sustained through persistent cytokine signalling, particularly via the IL-6–JAK pathway, and is reinforced by feed-forward loops involving RAGE, TLR4, CCL2, NF-κB, and the NLRP3 inflammasome.
This paper outlines the clinical consequences of persistent STAT3 dysregulation, linking it to malignancies, autoimmune disease, immunodeficiency, allergic responses, and metabolic exhaustion. New insights highlight STAT3’s role in astrocyte dysfunction, pericyte pathology, glymphatic impairment, and mitochondrial failure. Therapeutic strategies are reviewed, focusing on telmisartan, nicotinamide riboside, alpha-lipoic acid, low-dose naltrexone, hyperbaric oxygen, BPC-157, and GLP-1/GIP agonists.
Introduction
Long COVID, or Post-Acute Sequelae of COVID-19 (PASC), affects a significant proportion of individuals recovering from SARS-CoV-2 infection, with persistent symptoms such as fatigue, brain fog, and dysautonomia. Chronic inflammation and immune–metabolic reprogramming are increasingly recognized as central drivers.
STAT3 is a transcription factor regulating immune, inflammatory, and metabolic responses. In acute COVID-19, STAT3 hyperactivation contributes to cytokine storms, fibrosis, thrombosis, and immune dysfunction. In Long COVID, STAT3 remains persistently activated through interactions with RAGE, TLR4, CCL2, NF-κB, and NLRP3, creating a self-sustaining pathogenic network.
STAT3 Activation in Acute COVID-19
STAT3 is activated by cytokines such as IL-6 via the JAK pathway, leading to nuclear translocation and transcriptional activation. (Rezaei et al., 2021). In acute COVID-19, STAT3 is implicated in:
Hyper-inflammation: amplification of IL-6, TNF-α, and chemokines. (Rezaei et al., 2021).
Lung Fibrosis & Thrombosis: fibroblast proliferation, coagulation, endothelial dysfunction. (Rezaei et al., 2021).
Immune Dysfunction: lymphopenia, impaired NK-cell function, and immune exhaustion. (Rezaei et al., 2021).
These acute dynamics lay the foundation for its chronic role in Long COVID.
STAT3 in Long COVID: An Amplifier Within a Pathogenic Circuit
Persistent IL-6, DAMP-induced TLR4, RAGE activation, and NF-κB–CCL2 signalling maintain STAT3 in a phosphorylated state across monocytes, endothelial cells, and astrocytes. This generates several systemic consequences:
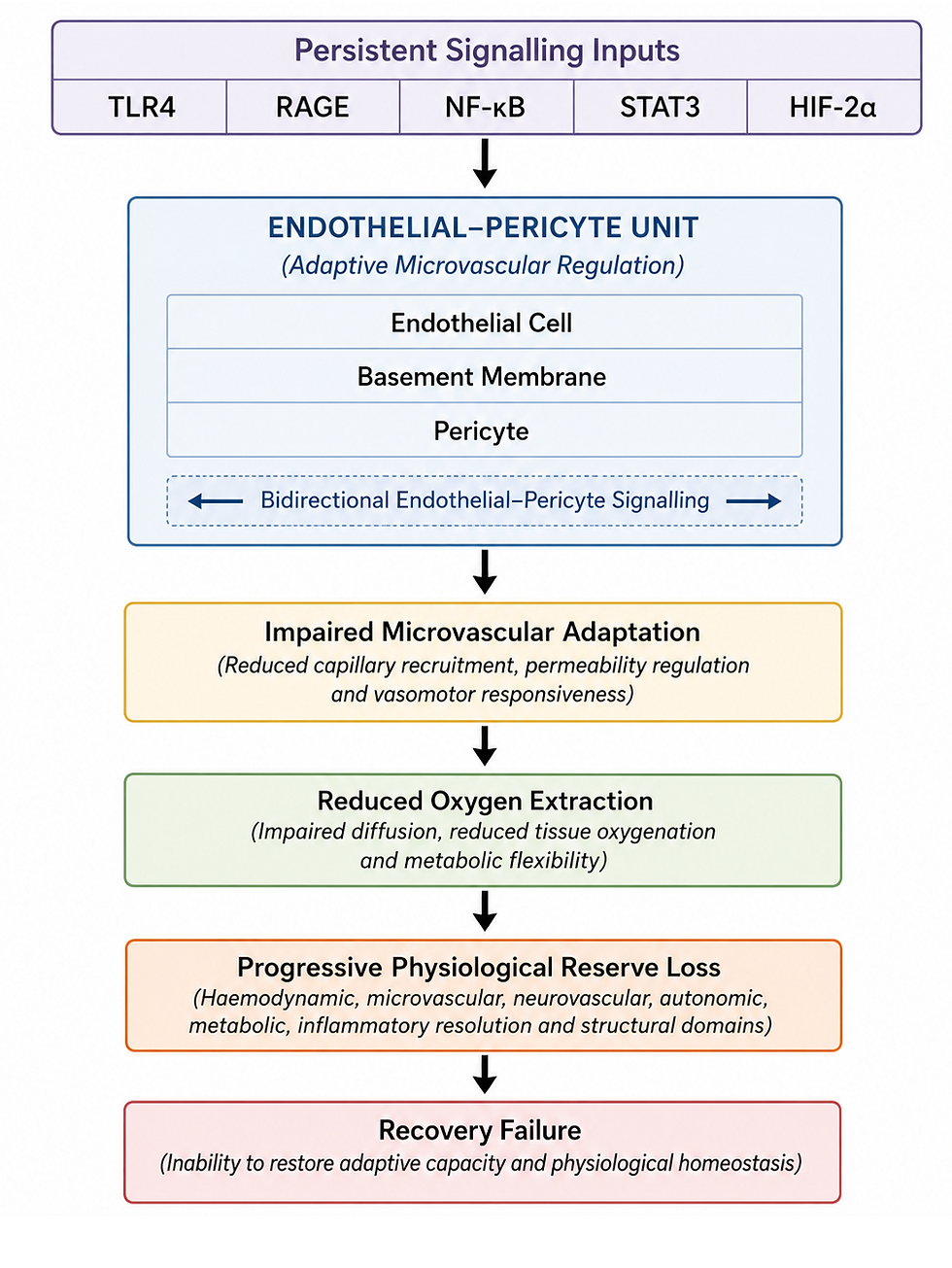
Astrocyte and Pericyte Dysfunction: STAT3 drives IL-6 secretion, disrupts aquaporin-4 polarity, and impairs glymphatic flow, producing neuroinflammation, cognitive decline, and upright head pressure.
NLRP3 Crosstalk: inflammasome activation stabilizes STAT3 via IL-1β and IL-18, perpetuating inflammation.
Mitochondrial Dysfunction: STAT3 suppresses mitophagy, inhibits PDH activity, and interferes with NAD+/SIRT3/SIRT4 signalling, underpinning post-exertional malaise and metabolic exhaustion.
Immune Exhaustion: chronic STAT3 signalling contributes to NK-cell dysfunction, impaired antiviral responses, and susceptibility to reactivation of latent viruses.
Thus, STAT3 is best understood not as an isolated driver but as a nodal amplifier within an interlocking immune–metabolic loop. (Deeks et al., 2022). (Douaud et al., 2022).( Wu et al., 2021). (Maucourant et al., 2021).
STAT3 and Malignancy
Persistent STAT3 activity promotes oncogenesis by enhancing proliferation, angiogenesis, and immune evasion. STAT3 also amplifies PD-L1 expression, suppressing T-cell responses. CCL2–STAT3 crosstalk further recruits tumour-associated macrophages, contributing to fibrosis and tumour microenvironment remodelling. These mechanisms reinforce the elevated malignancy risk seen in post-viral immune dysregulation.
Autoimmunity
STAT3 contributes to autoimmunity through Th17 differentiation and B-cell survival:
SLE: drives autoantibody production and nephritis.
RA: IL-6–STAT3 signalling in synoviocytes promotes joint destruction.
MS & IBD: STAT3–RORγt axis sustains pathogenic Th17 responses.
Genetic predispositions (e.g., STAT3 gain-of-function mutations, CCL2 and TLR4 variants) may amplify these autoimmune risks in Long COVID cohorts.
Immunodeficiency Syndromes
Loss-of-function STAT3: Hyper-IgE Syndrome with recurrent infections.
Gain-of-function STAT3: early-onset autoimmunity, lymphoproliferation, and immune exhaustion.
Long COVID phenocopies elements of these disorders, suggesting STAT3 imbalance—whether hypo- or hyper-functional—drives vulnerability to infection and autoimmunity alike.
Allergic and Mast Cell Disorders
STAT3 regulates mast cell exocytosis and IgE signalling. Hyperactivation amplifies allergic responses, contributing to mast cell activation syndrome (MCAS)-like symptoms and post-viral hypersensitivity states.
Molecular Pathways
IL-6–JAK–STAT3: core inflammatory driver.
IL-23–STAT3–RORγt: sustains Th17 autoimmunity.
PD-1/PD-L1–STAT3: immune evasion, antiviral suppression.
RAGE–TLR4–NF-κB–CCL2–STAT3 loop: interlocked innate-adaptive immune crosstalk.
NLRP3–STAT3 axis: chronic inflammasome stabilization and cytokine amplification.
Therapeutic Implications
Direct STAT3 inhibitors remain experimental, but indirect strategies are available:
Telmisartan: PPAR-γ agonism, RAGE suppression, endothelial protection. (Yamagishi et al 2008 )
Nicotinamide Riboside + Magnesium: NAD+ restoration, SIRT activation, metabolic recovery.
Alpha-Lipoic Acid (ALA): redox balance, PDH reactivation, glutamate modulation.
Low-Dose Naltrexone (LDN): TLR4 and microglial modulation.
Hyperbaric Oxygen (HBO): anti-hypoxic and anti-inflammatory.
BPC-157 and other peptides: endothelial, ECM, and mitochondrial repair, with indirect STAT3 modulation.(experimental)
GLP-1/GIP Agonists (e.g., tirzepatide): suppression of IL-6/STAT3 signalling, vascular and metabolic support.
These interventions may represent a therapeutic scaffold targeting STAT3-driven immune–metabolic dysfunction.
Conclusion
Persistent STAT3 activation in Long COVID is not an isolated abnormality but a nodal amplifier within a self-sustaining immune–metabolic loop involving RAGE, NF-κB, CCL2, and NLRP3. This integrated circuit drives malignancy risk, autoimmunity, immune deficiency, allergic hypersensitivity, vascular dysfunction, and metabolic collapse.
Targeting STAT3 directly remains a challenge, but modulating its supporting network with telmisartan, metabolic restoration agents, redox stabilizers, peptides, and GLP-1/GIP agonists provides a rational, multi-pronged therapeutic pathway. Understanding STAT3 within its broader immune–metabolic ecosystem may be the key to preventing chronic post-viral sequelae and improving patient outcomes.
References
1. Chiappalupi, S. et al. (2021). Targeting RAGE to prevent SARS-CoV-2-mediated multiple organ failure. Frontiers in Immunology, 12, 637542.
2. Deeks, S.G. et al. (2022). Long COVID (PASC) maintained by TLR4/RAGE-loop. International Journal of Molecular Sciences, 2(3), 33.
3. Douaud, G. et al. (2022). SARS-CoV-2 is associated with changes in brain structure in UK Biobank. Nature, 604(7907), 694–700.
4. Huang, C. et al. (2020). Unique immunological profile in patients with COVID-19. Cell, 182(1), 8–20.
5. Kaur, J. et al. (2021). Hyperactivated RAGE in comorbidities as a risk factor for severe COVID-19—The Role of RAGE-RAS Crosstalk. International Journal of Molecular Sciences, 22(12), 6494.
6. Liu, Y. et al. (2023). RAGE engagement by SARS-CoV-2 enables monocyte infection and underlies COVID-19 severity. Nature Immunology, 24(1), 105–115.
7. Maucourant, C. et al. (2021). NK cell dysfunction in patients with COVID-19. Nature Immunology, 22(10), 1252–1261.
8. Rezaei, N. et al. (2021). Contribution of STAT3 to the pathogenesis of COVID-19. Journal of Medical Virology, 93(5), 2759–2767.
9. Wu, H. et al. (2021). Activation of STAT3 signaling in kidney of COVID-19 patients. Infection, 49(6), 1231–1239.
10. Jones, S.A. et al. (2011). IL-6 signaling pathways in inflammatory diseases. The Journal of Clinical Investigation, 121(9), 3375–3383.
11. Smith, K.A. (2018). STAT3 activation in human diseases. Clinical Immunology, 192, 12–17.
12. O'Shea, J.J. et al. (2013). The JAK-STAT pathway: impact on human disease and therapeutic intervention. Annual Review of Medicine, 64, 311–328.
13. Durant, L. et al. (2010). Diverse targets of the transcription factor STAT3 contribute to T cell pathogenicity and homeostasis. Immunity, 32(5), 605–615.
14. Zhu, J. et al. (2010). Differentiation of effector CD4 T cell populations. Annual Review of Immunology, 28, 445–489.
15. Lovato, P. et al. (2011). STAT3 activation contributes to Th17 cell induction and skin inflammation in human psoriasis. Journal of Investigative Dermatology, 131(7), 1339–1348.
16. Yang, L., Xie, X., Tu, Z. et al. The signal pathways and treatment of cytokine storm in COVID-19. Sig Transduct Target Ther 6, 255 (2021). https://doi.org/10.1038/s41392-021-00679-0
17. Ahuja A, Kim E, Sung GH, Cho JY. STAT3 Differentially Regulates TLR4-Mediated Inflammatory Responses in Early or Late Phases. Int J Mol Sci. 2020;21(20):7675. Published 2020 Oct 16. doi:10.3390/ijms21207675
18. Xia C, Braunstein Z, Toomey AC, Zhong J, Rao X. S100 Proteins As an Important Regulator of Macrophage Inflammation. Front Immunol. 2018 Jan 5;8:1908. doi: 10.3389/fimmu.2017.01908. PMID: 29379499; PMCID: PMC5770888.
19. Bu LL, Yu GT, Wu L, et al. STAT3 Induces Immunosuppression by Upregulating PD-1/PD-L1 in HNSCC. J Dent Res. 2017;96(9):1027-1034. doi:10.1177/0022034517712435
20. Suzanne Ostrand-Rosenberg, Lucas A. Horn, Samuel T. Haile; The Programmed Death-1 Immune-Suppressive Pathway: Barrier to Antitumor Immunity. J Immunol 15 October 2014; 193 (8): 3835–3841. https://doi.org/10.4049/jimmunol.1401572
21. Chen WY, Chen H, Hamada K, et al. Transcriptomics identifies STAT3 as a key regulator of hippocampal gene expression and anhedonia during withdrawal from chronic alcohol exposure. Transl Psychiatry. 2021;11(1):298. Published 2021 May 20. doi:10.1038/s41398-021-01421-8
22. Li M, Plecitá-Hlavatá L, Dobrinskikh E, McKeon BA, Gandjeva A, Riddle S, Laux A, Prasad RR, Kumar S, Tuder RM, Zhang H, Hu CJ, Stenmark KR. SIRT3 Is a Critical Regulator of Mitochondrial Function of Fibroblasts in Pulmonary Hypertension. Am J Respir Cell Mol Biol. 2023 Nov;69(5):570-583. doi: 10.1165/rcmb.2022-0360OC. PMID: 37343939; PMCID: PMC10633840.
23. Miyamoto T, Mori T, Yoshimura A, Toyama T. STAT3 is critical to promote inflammatory cytokines and RANKL expression in inflammatory arthritis. Arthritis Res Ther. 2012;14(Suppl 1):P43. doi:10.1186/ar3644
24. Erlich TH, Yagil Z, Kay G, Peretz A, Migalovich-Sheikhet H, Tshori S, Nechushtan H, Levi-Schaffer F, Saada A, Razin E. Mitochondrial STAT3 plays a major role in IgE-antigen-mediated mast cell exocytosis. J Allergy Clin Immunol. 2014 Aug;134(2):460-9. doi: 10.1016/j.jaci.2013.12.1075. Epub 2014 Feb 28. PMID: 24582310.
25. Yamagishi S, Matsui T, Nakamura K, Takeuchi M, Inoue H. Telmisartan inhibits advanced glycation end products (AGEs)-elicited endothelial cell injury by suppressing AGE receptor (RAGE) expression via peroxisome proliferator-activated receptor-gammaactivation. Protein Pept Lett. 2008;15(8):850-3. doi: 10.2174/092986608785203746. PMID: 1885575
Comments